
A histologia (do grego: hydton = tecido + logos =
estudos) é a ciência que estuda os tecidos biológicos, desde a sua
formação (origem), estrutura (tipos diferenciados de células) e
funcionamento.
Mas o que é tecido?
O corpo de um organismo multicelular é
constituído por diferentes tipos de células, especializadas em realizar
diversas funções. As células com determinado tipo de especialização organizam-se em grupos, constituindo os tecidos.
Alguns tecidos são formados por células que possuem a mesma estrutura;
outros são formados por células que têm diferentes formas e funções, mas
que juntas colaboram na realização de uma função geral maior.
A diferenciação dos tecidos e a conquista do ambiente terrestre
Dentre as diversas adaptações que favoreceram a conquista do meio terrestre pelos vertebrados destacam-se um eficiente revestimento
corporal impermeabilizado, um adequado sistema esquelético de suporte
do organismo e de seus órgãos e um hábil mecanismo que permite a
movimentação do organismo pelo meio. No homem, essas três
tarefas são desempenhadas, na ordem, pela pele, pelo conjuntivo de ossos
do sistema esquelético e pelos inúmeros músculos componentes do sistema
muscular. Ossos e músculos constituem o sistema locomotor.
Como são formados os tecidos?
Todos os tecidos presentes nos vertebrados adultos são formados a partir de três tipos de folhetos germinativos: endoderma, ectoderma e mesoderma.
Cada um desses, durante o desenvolvimento embrionário, é responsável
por uma genealogia de células especializadas quanto à forma e função.

Os destinos finais (organogênese) desses folhetos germinativos, na formação dos tecidos e órgão humanos, são os seguintes.
Ectoderma
- Epiderme e anexos cutâneos (pêlos e glândulas mucosas);
- Todas as estruturas do sistema nervoso (encéfalo, nervos, gânglios nervosos e medula espinhal);
- Epitélio de revestimento das cavidades nasais, bucal e anal.
Mesoderma
- Forma a camada interna da pele (derme);
- Músculos lisos e esqueléticos;
- Sistema circulatório (coração, vasos sanguíneos, tecido linfático, tecido conjuntivo);
- Sistema esquelético (ossos e cartilagem);
- Sistema excretor e reprodutor (órgãos genitais, rins, uretra, bexiga e gônadas).
Endoderma
- Epitélio de revestimento e glândulas do trato digestivo, com exceção da cavidade oral e anal;
- Sistema respiratório (pulmão);
- Fígado e pâncreas.
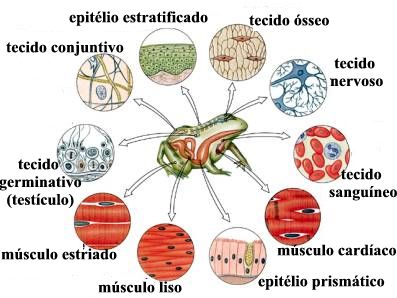
Tipos de Tecidos
Nos animais vertebrados há quatro grandes grupos de tecidos: o muscular, o nervoso, o conjuntivo (abrangendo também os tecidos ósseo, cartilaginoso e sanguíneo) e o epitelial, constituindo subtipos específicos que irão formar os órgãos e sistemas corporais.
Por exemplo: O sangue é considerado um tecido conjuntivo, com diversificadas células (as hemácias, os leucócitos e as plaquetas) e o plasma (água, sais minerais e diversas proteínas).
Por exemplo: O sangue é considerado um tecido conjuntivo, com diversificadas células (as hemácias, os leucócitos e as plaquetas) e o plasma (água, sais minerais e diversas proteínas).
Nos invertebrados estes tipos de tecido são
basicamente os mesmos, porém com organizações mais simples. A maioria
dos tecidos além de serem compostos de células, apresentam entre elas
substâncias intracelulares (intersticiais).

Especificação dos tecidos básicos
Epitélio → revestimento da superfície externa do corpo (pele), os órgãos (fígado, pulmão e rins) e as cavidades corporais internas;
Conjuntivo →
constituído por células e abundante matriz extracelulas, com função de
preenchimento, sustentação e transporte de substâncias;
Muscular → constituído por células com propriedades contráteis;
Nervoso → formado por células que constituem o sistema nervoso central e periférico (o cérebro, a medula espinhal e os nervos).
Tecido epitelial
A superfície externa do corpo e as cavidades corporais internas dos animais são revestidas por este tecido.
O tecido epitelial desempenha várias funções no organismo, como proteção do corpo (pele), absorção de substâncias úteis (epitélio do intestino) e percepção de sensações (pele), dependendo do órgão aonde se localizam.
Os tecidos epiteliais ou epitélios têm células perfeitamente justapostas, unidas por pequena quantidade de material cimentante, com pouquíssimo espaço intercelular.
Os epitélios não são vascularizados e não sangram quando feridos. A
nutrição das células se faz por difusão a partir dos capilares
existentes em outro tecido, o conjuntivo, adjacente ao epitélio a ele
ligado. O arranjo das células epiteliais pode ser comparado ao de
ladrilhos ou tijolos bem encaixados.

Os epitélios podem ser classificados quanto ao número de células:
- Quando os epitélios são formados por uma só camada de células, são chamados de epitélios simples ou uniestratificados (do latim uni, um, e stratum, camada).
- Já os epitélios formados por mais de uma camada de células são chamados estratificados.
- Existem ainda epitélios que, apesar de formados por uma única camada celular, têm células de diferentes alturas, o que dá a impressão de serem estratificados. Por isso, eles costumam ser denominados pseudo-estratificados.
Quanto à forma das células, os epitélios podem ser classificados em:
- Pavimentosos, quando as células são achatadas como ladrilhos;
- Cúbicos, quando as células tem forma de cubo, ou
- Prismáticos, quando as células são alongadas , em forma de coluna.
No epitélio que reveste a bexiga, a forma das
células é originalmente cúbica, mas elas se tornam achatadas quando
submetidas ao estiramento causado pela dilatação do órgão. Por isso,
esse tipo de epitélio é de denominado, por alguns autores, epitélio de
transição.
Os tecidos epiteliais, também chamados epitélios, são classificados em dois tipos principais: epitélios de revestimento e epitélios glandulares.
Epitélios de revestimento
Funciona como uma membrana que isola o organismo, ou parte dele, do meio externo.
Está relacionado ao revestimento e proteção de
superfícies externas (por exemplo, na pele) e internas (por exemplo, no
estômago). Atua, também, na absorção de substâncias, na secreção de
diversos produtos, na remoção de impurezas e pode conter vários tipos de
receptores sensoriais (notadamente na pele).
Pele: Órgão de contato
Nos vertebrados, a pele é importante órgão de
contato com o meio. A conquista do ambiente terrestre pelos vertebrados
tornou-se possível, entre outras coisas, a partir do isolamento e
proteção do corpo e de mecanismos de relação do ser vivo com o meio.
O tato, a visão, a olfação, a gustação e a
audição são úteis no relacionamento do animal com o ambiente. A pele,
órgão responsável pelas sensações táteis, apresenta diferentes tipos de
“sensores”, que registram e informam ao ser vivo variações de
temperatura (calor ou frio) e pressão (toques, choques, pancadas). A
pele é, ainda, importante órgão de defesa contra diversos tipos de
agentes infecciosos.
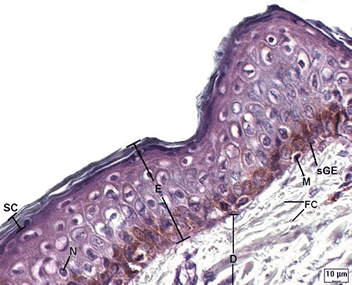
Tecido Epitelial de Revestimento Pluriestratificado Pavimentoso Queratinizado. Microscopia óptica. (E) Epiderme, (D) Derme, (SC) Células queratinizadas e cera.
Considerando o corpo inteiro, a pele de uma pessoa chega a pesar 5 Kg e tem uma área total de 18 m2. É, portanto o maior órgão do nosso corpo.
A histologia da pele
Nos mamíferos, a pele é órgão composto por duas camadas: epiderme e derme.
A epiderme é um tecido epitelial pluriestratificado.
É formada por estratos (ou camadas), dos quais destacam-se o estrato
basal (também chamado de estrato germinativo), que fica apoiado na derme
e é formado por células de aspecto cúbico. Nessa camada é intensa a
atividade de divisão celular mitótica, que repõe constantemente as
células perdidas no desgaste diário a que a superfície desse tecido está
sujeito. À medida que novas células são formadas, elas vão sendo
“empurradas” para formar as demais células, até ficarem expostas na
superfície da pele.
A derme é uma
camada formada por tecido conjuntivo do tipo denso, cujas fibras ficam
orientadas em diversas direções. Vários tipos de células são
encontrados, destacando-se os fibroblastos e os macrófagos. Nervos,
terminações nervosas, diferentes tipos de corpúsculos sensoriais e uma
ampla rede de capilares sanguíneos cruzam a derme em várias direções.
Ela é um importante tecido de manutenção e de apoio. Os nutrientes
existentes no sangue difundem-se para as células epidérmicas.
Nos mamíferos, a derme é atravessada por
finas faixas de células musculares, os músculos eretores dos pêlos, cuja
contração é involuntária e permite aumentar a camada de ar retirada
entre os pêlos, que contribui para o isolamento térmico. Mecanismo
semelhante ocorre nas aves, com as penas.
Abaixo da derme, há uma camada de tecido
conjuntivo frouxo, o tecido celular subcutâneo (também conhecido como
tela subcutânea e hipoderme), que não faz parte da pele, mas estabelece a
sua ligação com as estruturas adjacentes, permitindo o seu
deslizamento. Em determinadas regiões do corpo, a hipoderme contém um
número variável de camadas de células adiposas, formando o panículo
adiposo (o popular “toucinho de porco”), importante como reserva de
energia, isolante térmico e facilitador da flutuação na água.
Sensores da pele
Diversos tipos de estruturas sensoriais conferem à pele a função de relacionamento com o meio ambiente.
Distribuído por toda a pele, são basicamente dendritos de neurônios sensoriais (terminações
nervosas livres), sendo que alguns são envoltos por uma cápsula de
células conjuntivas ou epiteliais e, por isso, esses receptores são
capsulados.

Anexos da Pele
Três estruturas da pele, derivadas da epiderme, são extremamente importantes na adaptação dos mamíferos ao meio terrestre: pêlos,
que auxiliam no isolamento térmico; glândulas sudoríparas, que
desempenham o papel importante na regulação da temperatura corpórea; e
glândulas sebáceas, que lubrificam a pele e estruturas anexas.

A pele é um tipo de tecido epitelial chamado epitélio de revestimento.
Os tecidos epiteliais de revestimento são também encontrados revestindo
as cavidades corporais internas dos animais.
Epitélio de revestimento intestinal
O tecido que reveste internamente o intestino
delgado é um bom exemplo de epitélio especializado em absorver
nutrientes e permitir que eles passem da cavidade intestinal para o
sangue.
A alta capacidade de absorção do epitélio
intestinal se deve ao fato de suas células possuírem, na membrana a
borda livre (isto é, a borda voltada para a cavidade intestinal), muitas
projeções finas e alongadas, que lembrem dedos de uma luva, chamadas
microvilosidades.

Cálculos da área de membrana que constitui as
microvilosidades mostram que elas aumentam quinhentas vezes a área
superficial de cada célula, em comparação com a área de células que têm a
borda lisa. O mesmo tipo de cálculo nos leva a concluir que o intestino
delgado humano apresenta uma superfície de absorção de mais de 300 m2, equivalente à área de uma quadra de esportes de 20 m de comprimento por 15 m de lado.
A renovação das células epiteliais
A mitose é um processo
frequente nas células epiteliais, as quais têm vida curta e precisam ser
constantemente renovadas. A velocidade dessa renovação varia de
epitélio para epitélio. As células que se renovam mais rapidamente são
do epitélio intestinal: num prazo de 2 a 5 dias são substituídas por
células novas. As que se renovam mais lentamente são as células do
pâncreas que demoram 50 dias para serem substituídas.

Na pele, a renovação da epiderme ocorre em média a cada 30 dias. No
couro cabeludo, pode ocorrer uma disfunção em que a descamação de parte
da epiderme acontece a cada 3 ou 4 dias, formando as caspas.
Especialização das células epiteliais
As células dos tecidos epiteliais mantêm-se
aderidas umas às outras por meio de estruturas especializadas,
genericamente chamadas junções celulares.
Observe essas junções na figura abaixo.

Desmossomos
Uma das mais importantes junções celulares é o desmossomo (do grego desmos, ligação, e somatos,
corpo). Um desmossomo pode ser comparado a um botão de pressão
constituído por duas metades que se encaixam, estando uma metade
localizada na membrana de uma das células e a outra na célula vizinha.

Em cada célula existe uma placa circular de proteína, situada bem junto à
membrana. Das placas partem substâncias colantes, chamadas
desmogleínas, que atravessam as membranas e grudam as células na região
de contato. As placas também estão ligadas a um grande número de
filamentos constituídos da proteína queratina.


Zona de oclusão
Outro tipo de junção celular presente em
muitos epitélios é a zona de oclusão, uma espécie de cinturão adesivo
situado junto a borda livre das células epiteliais.
A zona de oclusão mantém as células vizinhas
tão encostadas que impede a passagem de moléculas entre elas. Assim,
substâncias eventualmente presentes em uma cavidade revestida por tecido
epitelial não podem penetrar no corpo, a não ser atravessando
diretamente as células.

Lâmina basal e hemidesmossomos
Sob um tecido epitelial há sempre uma espécie de
tapete de moléculas de proteínas ao qual as células se ligam: a lâmina
basal. As bases das células epiteliais ficam aderidas a lâmina basal por
meio de estruturas celulares especiais, denominadas hemidesmossomos.
Estes lembram desmossomos, mas possuem estrutura e função diferentes,
conectando as bases das células epiteliais à lamina basal, em vez de
ligarem as membranas de células vizinhas, como fazem os desmossomos.

Tecido epitelial glandular
As células do tecido epitelial glandular produzem substâncias chamadas secreções, que podem ser utilizadas e outras partes do corpo ou eliminadas do organismo. Essas secreções podem ser:
- mucosas, quando espessas e ricas em muco, Ex. glândulas salivares
- serosas, quando fluidas, aquosas, claras e ricas e proteínas. Ex. glândulas secretoras do pâncreas
- Podem também ser mistas, quando ocorrem secreções mucosas e serosas juntas. Ex. Glândulas salivares parótidas.
As glândulas podem ser unicelulares,
como a glândula caliciforme (que ocorre por exemplo, no epitélio da
traqueia), ou multicelulares, como a maioria das glândulas.

Em amarelo glândulas caliciformes do intestino.


Em verde a parte secretora de glândulas multicelulares exócrina e endócrina, respectivamente.
As glândulas multicelulares
originam-se sempre dos epitélios de revestimento, por proliferação de
suas células para o interior do tecido conjuntivo subjacente e posterior
diferenciação.
Três tipos de glândulas multicelulares
Glândulas exócrinas: apresentam
a porção secretora associada a dutos que lançam suas secreções para
fora do corpo (como as glândulas sudoríparas, lacrimais, mamárias e
sebáceas) ou para o interior de cavidades do corpo (como as glândulas
salivares);
Glândulas endócrinas:
não apresentam dutos associados à porção secretora. As secreções são
denominadas hormônios e lançadas diretamente nos vasos sanguíneos e
linfáticos. Exemplos, hipófise, glândulas da tireoide, glândulas
paratireódeas e glândulas adrenais;
Glândulas mistas:
apresentam regiões endócrinas e exócrinas ao mesmo tempo. É o caso do
pâncreas, cuja porção exócrina secreta enzimas digestivas que são
lançadas no duodeno, enquanto a porção endócrina é responsável pela
secreção dos hormônios insulina e glucagon. Esses hormônios atuam,
respectivamente, na redução e no aumento dos níveis de glicose no
sangue.

Tecido conjuntivo
Os tecidos conjuntivos têm origem mesodérmica.
Caracterizam-se morfologicamente por
apresentarem diversos tipos de células imersas em grande quantidade de
material extracelular, substância amorfa ou matriz, que é sintetizado
pelas próprias células do tecido.
A matriz é uma massa amorfa, de aspecto
gelatinoso e transparente. É constituída principalmente por água e
glicoproteínas e uma parte fibrosa, de natureza protéica, as fibras do
conjuntivo.
As células conjuntivas são de diversos tipos. As principais são apresentadas a seguir.
Fibroblasto
Célula metabolicamente ativa,
contendo longos e finos prolongamentos citoplasmáticos. Sintetiza o
colágeno e as substãncias da matriz (substância intercelular).

Macrófago
Célula ovoide, podendo conter
longos prolongamentos citoplasmáticos e inúmeros lisossomos.
Responsável pela fagocitose e pinocitose de pertículas estranhas ou não
ao organismo. Remove restos celulares e promove o primeiro combate aos
microrganismos invasores do nosso organismo. Ativo no processo de
involução fisiológica de alguns órgãos ou estrutura. É o caso do útero
que, após o parto, sofre uma redução de volume.

Mastócito
Célula globosa, grande, sem
prolongamentos e repleta de grânulos que dificultam, pela sua
quantidade, a visualização do núcleo. Os grânulos são constituídos de
heparina (substãncia anticoagulante) e histamina (substãncia envolvida
nos processos de alergia). Esta última substãncia é liberada em ocasiões
de penetração de certos antígenos no organismo e seu contato com os
mastócitos, desencadeando a consequênte reação alérgica.

Plasmócito
Célula ovoide, rica em
retículo endoplasmático rugoso (ou granular). Pouco numeroso no conjunto
normal, mas abundante em locais sujeitos à penetração de bactérias,
como intestino, pele e locais em que existem infecções crônicas.
Produtor de todos os anticorpos no combate a microorganismos. É
originado no tecido conjuntivo a partir da diferenciação de células
conhecidas como linfócitos B.

Os diferentes tipos de tecido conjuntivo estão
amplamente distribuídos pelo corpo, podendo desempenhar funções de
preenchimento de espaços entre órgãos, função de sustentação, função de
defesa e função de nutrição.
A classificação desses tecidos baseia-se na
composição de suas células e na proporção relativa entre os elementos da
matriz extracelular. Os principais tipos de tecidos conjuntivos são: frouxo, denso, adiposo, reticular ou hematopoiético, cartilaginoso e ósseo.
Tecido conjuntivo frouxo
O tecido conjuntivo frouxo preenche espaços não
ocupados por outros tecidos, apoia e nutre células epiteliais, envolve
nervos, músculos e vasos sanguíneos linfáticos.
Além disso, faz parte da estrutura de muitos órgãos e desempenha importante papel em processos de cicatrização.

É o tecido de maior distribuição no corpo humano.
Sua substância fundamental é viscosa e muito hidratada. Essa
viscosidade representa, de certa forma, uma barreira contra a penetração
de elementos estranhos no tecido. É constituído por três componentes
principais: células de vários tipos, três tipos de fibras e matriz.
Tipos de fibras
As fibras presentes no tecido conjuntivo frouxo são de três tipos: colágenas, elásticas e reticulares.
As fibras colágenas
são constituídas de colágeno, talvez a proteína mais abundante no reino
animal. São grossas e resistentes, distendendo-se pouco quando
tensionadas. As fibras colágenas presentes na derme conferem resistência
a nossa pele, evitando que elase rasgue, quando esticada.

As fibras elásticas são longos fios de uma
proteína chamada elastina. Elas conferem elasticidade ao tecido
conjuntivo frouxo, completando a resistência das fibras colágenas.
Quando você puxa e solta à pele da parte de cima da mão, são as fibras
elásticas que rapidamente devolvem à pele sua forma original. A perda da
elasticidade da pele, que ocorre com o envelhecimento, deve-se ao fato
de as fibras colágenas irem, com a idade, se unindo umas às outras,
tornando o tecido conjuntivo mais rígido.

As fibras reticulares são ramificadas e formam um trançado firme que liga o tecido conjuntivo aos tecidos vizinhos.

Tipos de células
O tecido conjuntivo frouxo contém dois principais de células: fibroblastos e macrófagos.
Os fibroblastos
têm forma estrelada núcleo grande. São eles que fabricam e secretam as
proteínas que constituem as fibras e a substância amorfa.
Os macrófagos
são grandes e ameboides, deslocando-se continuamente entre as fibras à
procura de bactérias e restos de células. Sua função é limpar o tecido,
fagocitando agentes infecciosos que penetram no corpo e, também, restos
de células mortas. Os macrófagos, alem disso identificam substâncias
potencialmente perigosas ao organismo, alertando o sistema de defesa do
corpo.

Outros tipos celulares presentes no tecido conjuntivo frouxo são as células mesenquimatosas e os plasmócitos.
As células mesenquimatosas são dotadas de alta capacidade de
multiplicação e permitem a regeneração do tecido conjuntivo, pois dão
origem a qualquer tipo de célula nele presente. Os plasmócitos são
células especializadas em produzir os anticorpos que combatem
substâncias estranhas que penetram no tecido.
Tecido conjuntivo denso
No tecido conjuntivo denso há predomínio de fibroblastos e fibras colágenas.
Dependendo do modo de organização dessas fibras, esse tecido pode ser classificado de diferentes formas, como veremos a seguir.

Classificações:
- não modelado: formado por fibras colágenas entrelaçadas, dispostas em feixes que não apresentam orientação fixa, o que confere resistência e elasticidade. Esse tecido forma as cápsulas envoltórias de diversos órgãos internos, e forma também um a derme, tecido conjuntivo da pele;

modelado: formado por fibras colágenas dispostas em
feixes com orientação fixa, dando ao tecido características de maior
resistência à tensão do que a dos tecidos não-modelados e frouxo; ocorre
nos tendões, que ligam os músculos aos ossos, e nos ligamentos, que
ligam os ossos entre si.

Tecido conjuntivo adiposo
Nesse tecido a substância intracelular é reduzida, e as células, ricas em lipídios, são denominadas células adiposas.
Ocorre principalmente sob a pele, exercendo
funções de reserva de energia, proteção contra choques mecânicos e
isolamento térmico. Ocorre também ao redor de alguns órgãos como os rins
e o coração.

As células adiposas possuem um grande vacúolo central de gordura, que
aumenta ou diminui, dependendo do metabolismo: se uma pessoa come pouco
ou gasta muita energia, a gordura das células adiposas diminui; caso
contrário, ela se acumula. O tecido adiposo atua como reserva de energia
para momentos de necessidade.

Tecido conjuntivo cartilaginoso
O tecido cartilaginoso, ou simplesmente cartilagem, apresentam consistência firme, mas não é rígido como o tecido ósseo.
Tem função de sustentação, reveste superfícies
articulares facilitando os movimentos e é fundamental para o crescimento
dos ossos longos.
Nas cartilagens não há nervos nem vasos
sanguíneos. A nutrição das células desse tecido é realizada por meio dos
vasos sanguíneos do tecido conjuntivo adjacente.
A cartilagem é encontrada no nariz, nos anéis da traqueia e dos brônquios, na orelha externa (pavilhão auditivo), na epiglote e em algumas partes da laringe.
Além disso, existem discos cartilaginosos entre as vértebras, que
amortecem o impacto dos movimentos sobre a coluna vertebral. No feto, o
tecido cartilaginoso é muito abundante, pois o esqueleto é inicialmente
formado por esse tecido, que depois é em grande parte substituído pelo
tecido ósseo.
O tecido cartilaginoso forma o esqueleto de
alguns animais vertebrados, como os cações, tubarões e raias, que são,
por isso, chamados de peixes cartilaginosos.
Há dois tipos de células nas cartilagens: os condroblastos (do grego chondros, cartilagem, e blastos,
“célula jovem”), que produzem as fibras colágenas e a matriz, com
consistência de borracha. Após a formação da cartilagem, a atividade dos
condroblastos diminui e eles sofrem uma pequena retração de volume,
quando passam a ser chamados de condrócitos (do grego chondros, cartilagem, e kytos,
célula). Cada condrócito fica encerrado no interior de uma lacuna
ligeiramente maior do que ele, moldada durante a deposição da matriz
intercelular.
As fibras presentes nesse tecido são as colágenas e as reticulares.

Legenda:
- Condroblasto
- Condrócito
- Grupo Isógeno
- Matriz Cartilaginosa
Tecido conjuntivo sanguíneo
O sangue (originado pelo tecido
hemocitopoiético) é um tecido altamente especializado, formado por
alguns tipos de células, que compõem a parte figurada, dispersas num
meio líquido – o plasma -, que corresponde à parte amorfa. Os
constituintes celulares são: glóbulos vermelhos (também denominados hemácias ou eritrócitos); glóbulos brancos (também chamados de leucócitos).
O plasma é composto principalmente de
água com diversas substâncias dissolvidas, que são transportadas
através dos vasos do corpo.

Todas as células do sangue são originadas na
medula óssea vermelha a partir das células indiferenciadas pluripotentes
(células-tronco). Como consequência do processo de diferenciação
celular, as células-filhas indiferenciadas assumem formas e funções
especializadas.
Plaquetas
Plaquetas são restos celulares originados da
fragmentação de células gigantes da medula óssea, conhecidas como
megacariócitos. Possuem substâncias ativas no processo de coagulação
sanguínea, sendo, por isso, também conhecidas como trombócitos (do
grego, thrombos = coágulo), que impedem a ocorrência de hemorragias.
Glóbulos vermelhos
Glóbulos vermelhos, hemácias ou eritrócitos (do grego, eruthrós = vermelho, e kútos
= célula) são anucleados, possuem aspecto de disco bicôncavo e diâmetro
de cerca de 7,2 m m. São ricos em hemoglobina, a proteína responsável
pelo transporte de oxigênio, a importante função desempenhada pelas
hemácias.
Glóbulos brancos
Glóbulos brancos, também chamados de
leucócitos (do grego, leukós = branco), são células sanguíneas
envolvidas com a defesa do organismo.
Essa atividade pode ser exercida por fagocitose ou por meio da produção de proteínas de defesa, os anticorpos.
Costuma-se classificar os glóbulos brancos
de acordo com a presença ou ausência, em seu citoplasma, de grânulos
específicos e agranulócitos, os que não contêm granulações específicas,
comuns a qualquer célula.

Tecido conjuntivo ósseo
O tecido ósseo tem a função de sustentação e ocorre nos ossos do esqueleto dos vertebrados.
É um tecido rígido graças à presença de matriz rica em sais de cálcio, fósforo e magnésio. Além desses elementos, a matriz é rica em fibras colágenas, que fornecem certa flexibilidade ao osso.
Os ossos são órgãos ricos em vasos
sanguíneos. Além do tecido ósseo, apresentam outros tipos de tecido:
reticular, adiposo, nervoso e cartilaginoso.
Por serem um estrutura inervada e irrigada, os ossos apresentam sensibilidade, alto metabolismo e capacidade de regeneração.
Quando um osso é serrado, percebe-se que ele é formado por duas partes: uma sem cavidades, chamada osso compacto, e outra com muitas cavidades que se comunicam, chamada osso esponjoso.

Essa classificação é de ordem macroscópica, pois
quando essas partes são observadas no microscópio nota-se que ambas são
formadas pela mesma estrutura histológica. A estrutura microscópica de
um osso consiste de inúmeras unidades, chamadas sistemas de Havers.
Cada sistema apresenta camadas concêntricas de matriz mineralizada,
depositadas ao redor de um canal central onde existem vasos sanguíneos e
nervos que servem o osso.
Os canais de Havers
comunicam-se entre si, com a cavidade medular e com a superfície externa
do osso por meio de canais transversais ou oblíquos, chamados canais
perfurantes (canais de Volkmann). O interior dos ossos é preenchido pela
medula óssea, que pode ser de dois tipos: amarela, constituída por
tecido adiposo, e vermelha, formadora de células do sangue.

Tipos de células do osso
As células ósseas ficam localizadas em pequenas cavidades existentes nas camadas concêntricas de matriz mineralizada.
Quando jovens, elas são chamadas osteoblastos (do grego osteon, osso, e blastos,
“célula jovem”) e apresentam longas projeções citoplasmáticas, que
tocam os osteoblastos vizinhos. Ao secretarem a matriz intercelular ao
seu redor, os osteoblastos ficam presos dentro de pequenas câmeras, das
quais partem canais que contêm as projeções citoplasmáticas.
Quando a célula óssea se torna madura, transforma-se em osteócito (do grego osteon, osso, e kyton,
célula), e seus prolongamentos citoplasmáticos se retraem, de forma que
ela passa a ocupar apenas a lacuna central. Os canalículos onde ficavam
os prolongamentos servem de comunicação entre uma lacuna e outra, e é
através deles que as substâncias nutritivas e o gás oxigênio
provenientes do sangue até as células ósseas.

Além dos osteoblastos e dos osteócitos, existem outras células
importantes no tecido ósseo: os osteoclástos (do grego klastos, quebrar,
destruir). Essas células são especialmente ativas na destruição de
áreas lesadas ou envelhecidas do osso, abrindo caminho para a
regeneração do tecido pelos osteoblastos. Os cientistas acreditam que os
ossos estejam em contínua remodelação, pela atividade conjunta de
destruição e reconstrução empreendidas, respectivamente, pelos
osteoclastos e osteoblastos. Você encontrará mais informações sobre os
osteoclastos no texto sobre remodelação óssea.

A formação do tecido ósseo
A ossificação - formação de tecido ósseo - pode se dar por dois processos: ossificação intramenbranosa e ossificação endocondral.
No primeiro caso, o tecido ósseo surge aos poucos em uma membrana de natureza conjuntiva, não cartilaginosa.
Na ossificação endocondral, uma peça de
cartilagem, com formato de osso, serve de molde para a confecção de
tecido ósseo. Nesse caso, a cartilagem é gradualmente destruída e
substituída por tecido ósseo.

Crescimento nos ossos longos
A ossificação endocondral ocorre na formação de ossos longos, como os das pernas e os dos braços.
Nesses ossos, duas regiões principais
sofrerão a ossificação: o cilindro longo, conhecido como diáfise e as
extremidades dilatadas, que correspondem as epífises.
Entre a epífise de cada extremidade e a
diáfise é mantida uma região de cartilagem, conhecida como cartilagem de
crescimento, que possibilitará a ocorrência constante de ossificação
endocondral, levando à formação de mais osso. Nesse processo, os
osteoclastos desempenham papel importante. Eles efetuam constantemente a
reabsorção de tecido ósseo, enquanto novo tecido ósseo é formado.

Os osteoclastos atuam como verdadeiros
demolidores de osso, enquanto os osteoblastos exercem papel de
construtores de mais osso. Nesse sentido, o processo de crescimento de
um osso depende da ação conjunta de reabsorção de osso preexistente e da
deposição de novo tecido ósseo. Considerando, por exemplo, o aumento de
diâmentro de um osso longo, é preciso efetuar a reabsorção de camada
interna da parede óssea, enquanto na parede externa deve ocorrer
deposição de mais osso.
O crescimento ocorre até que se atinja
determinada idade, a partir da qual a cartilagem de crescimento também
sofre ossificação e o crescimento do osso em comprimento cessa.
Remodelação óssea
Depois que o osso atinge seu tamanho e forma
adultos, o tecido ósseo antigo é constantemente destruído e um novo
tecido é formado em seu lugar, em um processo conhecido como
remodelação.
A remodelação ocorre em diferentes
velocidades nas várias partes do corpo. Por exemplo, a porção distal do
fêmur é substituída a cada 4 meses; já os ossos da mão são completamente
substituídos durante a vida inteira do indivíduo. A remodelação permite
que os tecidos já gastos ou que tenham sofrido lesões sejam trocados
por tecidos novos e sadios. Ela também permite que o osso sirva como
reserva de cálcio para o corpo.
Em um adulto saudável, uma delicada
homeostase (equilíbrio) é mantida entre a ação dos osteoclastos
(reabsorção) durante a remoção de cálcio e a dos osteoblastos (aposição)
durante a deposição de cálcio. Se muito cálcio for depositado, podem se
formar calos ósseos ou esporas, causando interferências nos movimentos.
Se muito cálcio for retirado, há o enfraquecimento dos ossos,
tornando-os flexíveis e sujeitos a fraturas.

O crescimento e a remodelação normais dependem de vários fatores
- suficientes quantidades de cálcio e fósforo devem estar presentes na dieta alimentar do indivíduo;
- deve-se obter suficiente quantidade de vitaminas, principalmente vitamina D, que participa na absorção do cálcio ingerido;
- o corpo precisa produzir os hormônios responsáveis pela atividade do tecido ósseo:
- Hormônio de crescimento (somatotrofina): secretado pela hipófise, é responsável pelo crescimento dos ossos;
- Calcitonina: produzida pela tireoide, inibe a atividade osteoclástica e acelera a absorção de cálcio pelos ossos;
- Paratormônio: sintetizado pelas paratireoides, aumenta a atividade e o número de osteoclastos, elevando a taxa de cálcio na corrente sanguínea;
- Hormônios sexuais: também estão envolvidos nesse processo, ajudando na atividade osteoblástica e promovendo o crescimento de novo tecido ósseo.
Com o envelhecimento, o sistema esquelético
sofre a perda de cálcio. Ela começa geralmente aos 40 anos nas mulheres e
continua até que 30% do cálcio nos ossos seja perdido, por volta dos 70
anos. Nos homens, a perda não ocorre antes dos 60 anos. Essa condição é
conhecida como osteoporose.
Outro efeito do envelhecimento é a redução
da síntese de proteínas, o que diminui a produção da parte orgânica da
matriz óssea. Como consequência, há um acúmulo de parte inorgânica da
matriz. Em alguns indivíduos idosos, esse processo causa uma
fragilização dos ossos, que se tornam mais susceptíveis a fraturas.
O uso de aparelhos ortodônticos é um exemplo
de remodelação dos ossos, neste caso, resultando na remodelação da
arcada dentária.
Os aparelhos exercem forças diferentes
daquelas a que os dentes estão naturalmente submetidos. Nos pontos em
que há pressão ocorre reabsorção óssea, enquanto no lado oposta há
deposição de matriz. Assim, os dentes movem-se pelos ossos da arcada
dentária e passam a ocupar a posição desejada.
Tecidos musculares
Os tecidos musculares são de origem mesodérmica e
relacionam-se com a locomoção e outros movimentos do corpo, como a
contração dos órgãos do tubo digestório, do coração e das artérias.
As células dos tecidos musculares são alongadas e recebem o nome de fibras musculares ou miócitos. Em seu citoplasma, são ricas em dois tipos de filamento protéico: os de actina e os de miosina, responsáveis pela grande capacidade de contração e distensão dessas células.
Quando um músculo é estimulado a se
contrair, os filamentos de actina deslizam entre os filamentos de
miosina. A célula diminui em tamanho, caracterizando a contração.
Tipos de tecido muscular
Há três tipos de tecido muscular: estriado esquelético, estriado cardíaco e liso. Cada um deles tem características próprias, adequadas ao papel que desempenham no organismo.

Tecido muscular estriado esquelético
O tecido muscular estriado esquelético constitui a maior parte da musculatura do corpo dos vertebrados, formando o que se chama popularmente de carne.
Essa musculatura recobre totalmente o
esqueleto e está presa aos ossos, daí ser chamada de esquelética. Esse
tipo de tecido apresenta contração voluntária (que depende da vontade do indivíduo).
Um músculo esquelético é um pacote de longas
fibras. Cada uma delas é uma célula dotada de muitos núcleos, chamado
miócitos multinucleados. Um fibra muscular pode medir vários centímetros
de comprimento, por 50 mm de espessura.

A célula muscular estriada apresenta, no seu citoplasma, pacotes de finíssimas fibras contráteis, as miofibrilas, dispostas longitudinalmente. Cada miofibrila corresponde a um conjunto de dois tipos principais de proteínas: as miosina, espessas, e as actinas,
finas. Esses proteínas estão organizados de tal modo que originam
bandas transversais, claras e escuras, características das células
musculares estriadas, tanto as esqueléticas como as cardíacas.

Os filamentos de miosina formam bandas escuras, chamadas anisotrópicas (banda A), e os de actina, bandas claras, chamadas isotrópicas (banda I).
No centro de cada banda I aparece uma linha mais escura, chamada linha Z.
O intervalo entre duas linhas Z consecutivas constitui um miômetro ou
sarcômero e correspondem à unidade contrátil da célula muscular.
No centro de cada banda A existe uma faixa mais clara, chamada banda H, bem visível nas células musculares relaxadas e que vai desaparecendo à medida que a contração muscular ocorre.

Na contração muscular, os miofilamentos não
diminuem de tamanho, mas os sarcômeros ficam mais curtos e toda a célula
muscular se contrai.
O encurtamento dos sarcômeros ocorre em
função do deslizamento dos miofilamentos finos sobre os grosso, havendo
maior sobreposição entre eles: a banda I diminui de tamanho, pois os
filamentos de actina deslizam sobre os de miosina, penetram na banda A e
reduzem a largura da banda H.
A membrana plasmática da célula muscular estriada esquelética costuma ser chamada sarcolema (do grego, sarcos, carne).

Na contração muscular, os miofilamentos não
diminuem de tamanho, mas os sarcômeros ficam mais curtos e toda a célula
muscular se contrai.
O encurtamento dos sarcômeros ocorre em
função do deslizamento dos miofilamentos finos sobre os grosso, havendo
maior sobreposição entre eles: a banda I diminui de tamanho, pois os
filamentos de actina deslizam sobre os de miosina, penetram na banda A e
reduzem a largura da banda H.
A membrana plasmática da célula muscular estriada esquelética costuma ser chamada sarcolema (do grego, sarcos, carne).
Tecido muscular estriado cardíaco
Apresenta miócitos estriados com um ou dois
núcleos centrais. Esse tecido ocorre apenas no coração e apresenta
contração independente da vontade do indivíduo (contração involuntária).
No músculo cardíaco essa contração é vigorosa e rítmica.

Essas células musculares são menores e
ramificadas, intimamente unidas entre si por estruturas especializadas e
típicas da musculatura cardíaca: os discos intercalares, que fazem a
conexão elétrica entre todas as células do coração. Assim, se uma célula
receber um estímulo suficientemente forte, ele é transmitido a todas as
outras células e o coração como um todo se contrai.
Essa transmissão do estímulo é feita por canais
de passagem de água e íons entre as células, que facilita a difusão do
sinal iônico entre uma célula e outra, determinando a onda rítmica de
contração das células. Os discos intercalares possuem estruturas de
adesão entre células que as mantêm unidas mesmo durante o vigoroso
processo de contração da musculatura cardíaca.

As células musculares cardíacas são capazes de
auto-estimulação, não dependendo de um estímulo nervoso para iniciar a
contração. As contrações rítmicas do coração são geradas e conduzidas
por uma rede de células musculares cardíacas modificadas que se
localizam logo abaixo do endocárdio, tecido que reveste internamente o
coração.
Existem numerosas terminações nervosas no
coração, mas o sistema nervoso atua apenas regulando o ritmo cardíaco às
necessidades do organismo.
Tecido muscular liso ou não-estriado
As células musculares lisas não apresentam
estriação transversal, característica das células musculares
esqueléticas e cardíacas.
A razão disso é que os filamentos de actina e
miosina não se encontram alinhados ao longo do comprimento da célula.
Acredita-se que eles estejam arranjados em espiral dentro da fibra
muscular lisa.

Os miócitos se apresentam uninucleados e fusiformes, isto é, alongadas e
coam as extremidades afiladas. Nessas células a contração é
involuntária e lenta. Você pode decidir quando lavar as suas mãos, mas
não controla conscientemente os movimentos de seu estômago ou a
contração de seu coração. Ocorre nas artérias, sendo responsável por sua
contração; ocorre também no esôfago, no estômago e nos intestinos,
sendo responsável pelo peristaltismo (ou peristalse) nesses órgãos. Os
movimentos peristálticos são contrações em ondas que deslocam o material
alimentar dentro desses órgãos do sistema digestório.

Tecido nervoso
Os seres vivos reagem aos estímulos ambientais.
Mudanças nas condições do ambiente, tais como sons, choques, calor e
frio, são percebidas pelo organismo, que reage adotando uma postura
correspondente ao estímulo.
Embora sejam os músculos que respondem aos
estímulos, é o tecido nervoso o responsável por sua recepção e escolha
da resposta adequada.
O tecido nervoso tem origem ectodérmica,
nele a substância intercelular praticamente não existe. Os principais
componentes celulares são os neurônios e as células da glia.
As células da glia ou neuroglia são vários tipos celulares relacionados com a sustentação e a nutrição dos neurônios, com a produção de mielina e com a fagocitose.
Os neurônios, ou células nervosas, têm a
propriedade de receber e transmitir estímulos nervosos, permitindo ao
organismo responder a alteração do meio. Os neurônios são alongados,
podendo atingir, em alguns casos, cerca de 1 metro de comprimento, como
nos neurônios que se estendem desde nossas costas até o pé. São células
formadas por um corpo celular ou pericário, de onde partem dois tipos de
prolongamento: dendritos e axônio.
Os dentritos são
prolongamentos ramificados da célula especializados em receber
estímulos, que também podem ser recebidos pelo corpo celular. O impulso
nervoso é sempre transmitido no sentido dendrito – corpo – axônio.

O axônio é uma expansão celular fina, alongada e de
diâmetro constante, com ramificações em sua porção final, de modo que o
impulso pode ser transmitido simultaneamente a vários destinos. É uma
estrutura especializada na transmissão de impulsos nervosos para outros
neurônios ou para outros tipos celulares, como as células de órgãos
efetores (musculares e glandulares).
Transmissão do impulso nervoso
Em um neurônio, os estímulos se propagam sempre
no mesmo sentido: são recebidos pelos dendritos, seguem pelo corpo
celular, percorrem o axônio e, da extremidade deste, são passados à
célula seguinte (dendrito – corpo celular – axônio).
O impulso nervoso que se propaga através do
neurônio é de origem elétrica e resulta de alterações nas cargas
elétricas das superfícies externa e interna da membrana celular.
A membrana de um neurônio em repouso
apresenta-se com carga elétrica positiva do lado externo (voltado para
fora da célula) e negativa do lado interno (em contato com o citoplasma
da célula). Quando essa membrana se encontra em tal situação, diz-se que
está polarizada. Essa diferença de cargas elétricas é mantida pela
bomba de sódio e potássio. Assim separadas, as cargas elétricas
estabelecem uma energia elétrica potencial através da membrana: o
potencial de membrana ou potencial de repouso (diferença entre as cargas
elétricas através da membrana).

Quando um estímulo químico, mecânico ou elétrico
chega ao neurônio, pode ocorrera alteração da permeabilidade da
membrana, permitindo grande entrada de sódio na célula e pequena saída
de potássio dela. Com isso, ocorre uma inversão das cargas ao redor
dessa membrana, que fica despolarizada gerando um potencial de ação.
Essa despolarização propaga-se pelo neurônio caracterizando o impulso
nervoso.
Imediatamente após a passagem do impulso, a
membrana sofre repolarização, recuperando seu estado de repouso, e a
transmissão do impulso cessa.
O estímulo que gera o impulso nervoso deve
ser forte o suficiente, acima de determinado valor crítico, que varia
entre os diferentes tipos de neurônios, para induzir a despolarização
que transforma o potencial de repouso em potencial de ação. Esse é o
estímulo limiar. Abaixo desse valor o estímulo só provoca alterações
locais na membrana, que logo cessam e não desencadeiam o impulso
nervoso.
Qualquer estímulo acima do limiar gera o
mesmo potencial de ação que é transmitido ao longo do neurônio. Assim,
não existe variação de intensidade de um impulso nervoso em função do
aumento do estímulo; o neurônio obedece à regra do “tudo ou nada”.
Dessa forma, a intensidade das sensações vai
depender do número de neurônios despolarizados e da frequência de
impulsos. Imagine uma queimadura no dedo. Quanto maior a área queimada,
maior a dor, pois mais receptores serão estimulados e mais neurônios
serão despolarizados.

A transmissão do impulso nervoso de um neurônio a
outro ou às células de órgãos efetores é realizada por meio de uma
região de ligação especializada denominada sinapse.
O tipo mais comum de sinapse é a química, em que
as membranas de duas células ficam separadas por um espaço chamado
fenda sináptica.
Na porção terminal do axônio, o impulso
nervoso proporciona a liberação das vesículas que contêm mediadores
químicos, denominados neuro-transmissores. Os mais comuns são
acetilcolina e adrenalina.
Esses neurotransmissores caem na fenda
sináptica e dão origem ao impulsos nervosos na célula seguinte. Logo a
seguir, os neurotransmissores que estão na fenda sináptica são
degradados por enzimas específicas, cessando seus efeitos.

Transmissão do impulso nervoso
Em um neurônio, os estímulos se propagam sempre
no mesmo sentido: são recebidos pelos dendritos, seguem pelo corpo
celular, percorrem o axônio e, da extremidade deste, são passados à
célula seguinte (dendrito – corpo celular – axônio).
O impulso nervoso que se propaga através do
neurônio é de origem elétrica e resulta de alterações nas cargas
elétricas das superfícies externa e interna da membrana celular.
A membrana de um neurônio em repouso
apresenta-se com carga elétrica positiva do lado externo (voltado para
fora da célula) e negativa do lado interno (em contato com o citoplasma
da célula). Quando essa membrana se encontra em tal situação, diz-se que
está polarizada. Essa diferença de cargas elétricas é mantida pela
bomba de sódio e potássio. Assim separadas, as cargas elétricas
estabelecem uma energia elétrica potencial através da membrana: o
potencial de membrana ou potencial de repouso (diferença entre as cargas
elétricas através da membrana).
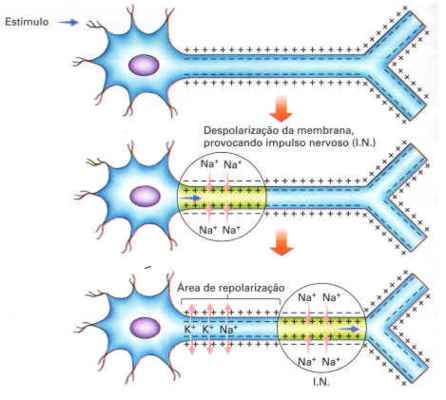
Quando um estímulo químico, mecânico ou
elétrico chega ao neurônio, pode ocorrera alteração da permeabilidade da
membrana, permitindo grande entrada de sódio na célula e pequena saída
de potássio dela. Com isso, ocorre uma inversão das cargas ao redor
dessa membrana, que fica despolarizada gerando um potencial de ação.
Essa despolarização propaga-se pelo neurônio caracterizando o impulso
nervoso.
Imediatamente após a passagem do impulso, a
membrana sofre repolarização, recuperando seu estado de repouso, e a
transmissão do impulso cessa.
O estímulo que gera o impulso nervoso deve
ser forte o suficiente, acima de determinado valor crítico, que varia
entre os diferentes tipos de neurônios, para induzir a despolarização
que transforma o potencial de repouso em potencial de ação. Esse é o
estímulo limiar. Abaixo desse valor o estímulo só provoca alterações
locais na membrana, que logo cessam e não desencadeiam o impulso
nervoso.
Qualquer estímulo acima do limiar gera o
mesmo potencial de ação que é transmitido ao longo do neurônio. Assim,
não existe variação de intensidade de um impulso nervoso em função do
aumento do estímulo; o neurônio obedece à regra do “tudo ou nada”.
Dessa forma, a intensidade das sensações vai
depender do número de neurônios despolarizados e da frequência de
impulsos. Imagine uma queimadura no dedo. Quanto maior a área queimada,
maior a dor, pois mais receptores serão estimulados e mais neurônios
serão despolarizados.
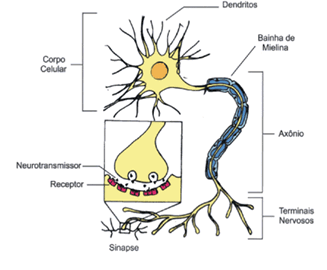
A transmissão do impulso nervoso de um
neurônio a outro ou às células de órgãos efetores é realizada por meio
de uma região de ligação especializada denominada sinapse.
O tipo mais comum de sinapse é a química, em que
as membranas de duas células ficam separadas por um espaço chamado
fenda sináptica.
Na porção terminal do axônio, o impulso
nervoso proporciona a liberação das vesículas que contêm mediadores
químicos, denominados neuro-transmissores. Os mais comuns são
acetilcolina e adrenalina.
Esses neurotransmissores caem na fenda
sináptica e dão origem ao impulsos nervosos na célula seguinte. Logo a
seguir, os neurotransmissores que estão na fenda sináptica são
degradados por enzimas específicas, cessando seus efeitos.
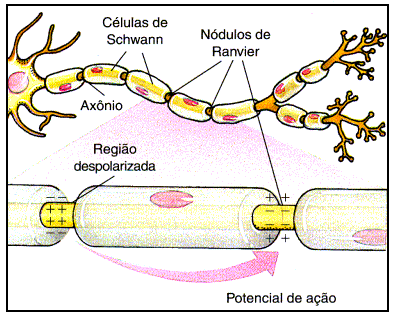
No sistema nervoso, verifica-se que os
neurônios dispõem-se diferenciadamente de modo a dar origem a duas
regiões com coloração distinta entre si e que podem ser notadas
macroscopicamente: a substância cinzenta, onde estão os corpos
celulares, e a substância branca, onde estão os axônios. No encéfalo
(com exceção do bulbo) a substância cinzenta está localizada
externamente em relação a substância branca, e na medula espinha e no
bulbo ocorre o inverso.
Os nervos são conjuntos de fibras nervosas organizadas em feixes, unidos por tecidos conjuntivo denso.
Regeneração das fibras nervosas
Assim com as células musculares do coração, os neurônios não se dividem mais depois de diferenciados.
Desse modo, se forem destruídos, não são mais
repostos. No entanto, os prolongamentos dos neurônios podem, dentro de
certos limites, sofrer regeneração, desde que o corpo celular não tenha
sido destruído.
Quando um axônio é cortado acidentalmente, o
que ocorre no caso de ferimentos na pele, a região que fica ligada ao
corpo celular é chamada coto proximal, e a que fica separada é chamada
coto distal. Este último degenera e é fagocitado pelos macrófagos, que
limpam a região lesada. Já o coto próxima cresce e se ramifica. Ao mesmo
tempo, células que formam a bainha de mielina do coto distal
modificam-se e proliferam, originando colunas celulares que servirão de
guia para os ramos que estão crescendo a partir do coto proximal. Quando
um desses ramos penetra nessa coluna de células, ele regenera
completamente o axônio.

Quando o espaço entre o coto proximal e o
distal é muito grande ou quando ocorre uma amputação, os ramos do coto
proximal crescem desordenadamente, entrelaçam-se e formam uma estrutura
muito sensível à dor, chamada neuroma de amputação.
Se gostou compartilha, se tem alguma dúvida ou sugestão me envia !!
Nenhum comentário:
Postar um comentário